Vol. 24, núm. 4 julio-agosto 2023
Los hidroides: pequeños grandes viajeros
Cecilia Odette Carral Murrieta, Elisa Serviere Zaragoza, Alejandra Mazariegos Villarreal y María A. Mendoza-Becerril CitaResumen
En este artículo descubrirás el fascinante mundo de los hidroides, criaturas acuáticas con ciclos de vida únicos. Estos animales pertenecen al filo Cnidaria, grupo taxonómico dentro del cual encontramos corales, anémonas, medusas y sus pólipos. Una de sus características más sorprendentes es su habilidad para viajar largas distancias, gracias a una estructura llamada hidrorriza, que les permite fijarse a diversos sustratos, incluyendo algas y objetos flotantes. A medida que exploramos la vida de los hidroides, entenderemos cómo su presencia puede afectar a las especies nativas y a actividades humanas como la acuicultura y la infraestructura marina. Además, conoceremos los esfuerzos para controlar la propagación de hidroides invasores. Aunque hemos realizado avances en su estudio, aún hay mucho por descubrir sobre estos organismos acuáticos y su papel en los ecosistemas marinos. Prepárate para sumergirte en el mundo fascinante de los hidroides y desentrañar sus secretos
Palabras clave: cnidarios, dispersión, hidroides, macroalgas, pólipos.
Hydroids: Those little big wanderers
Abstract
In this article, you will discover the fascinating world of hydroids, aquatic creatures with unique life cycles. These animals are part of the phylum Cnidaria, a taxonomic group within which we find corals, anemones, jellyfish, and polyps. One of their most surprising characteristics is their ability to travel long distances, thanks to a structure known as hydrorhiza, which allows them to attach to various substrates, including algae and floating objects. As we explore the life of hydroids, we will understand how their presence can affect native species and human activities such as aquaculture and marine infrastructure. In addition, we will learn about the efforts to control the spreading of invasive hydroids. Although we have made progress in their study, there is still much to discover about these aquatic organisms and their role in marine ecosystems. Get ready to immerse yourself in the fascinating world of hydroids and unravel their secrets.
Keywords: cnidarians, dispersion, hydroids, macroalgae, polyps.
¿Qué son los hidroides?
Los hidroides son animales acuáticos pertenecientes a un grupo taxonómico conocido como Cnidaria, dentro del cual encontramos unas 3761 especies (WoRMS, 2023), las cuales comparten un plan básico de organización e incluyen corales, medusas y sus pólipos. Dentro de los cnidarios se encuentra un grupo llamado clase Hydrozoa, cuyo nombre proviene del griego hydra, serpiente acuática, y zoon, animal. Una característica interesante de los hidrozoos es que tienen ciclos de vida con alternancia de generaciones, lo que significa que podemos encontrarnos dos etapas de vida diferentes dentro de una misma especie: una etapa de pólipo y una etapa de medusa (Mills et al., 2007). A la etapa pólipo se le conoce como hidroide, es sésil y bentónica, es decir, vive fija a sustratos, mientras que la etapa medusa, conocida como hidromedusa, generalmente es planctónica y móvil, por lo que se mantiene flotando en la columna de agua (Mills et al., 2007). Sin embargo, en varias especies de hidrozoos una de las dos fases nunca se presenta en el ciclo de vida, por ejemplo, hay especies de pólipos que jamás producen medusas (Mills et al., 2007; ver figura 1). Tanto los hidroides como las hidromedusas producen larvas plánulas, que son muy importantes en la dispersión, puesto que nadan por cortos períodos de tiempo hasta fijarse a un sustrato y dar lugar a un nuevo hidroide o por el contrario se convierten en una hidromedusa (Sommer, 1992).
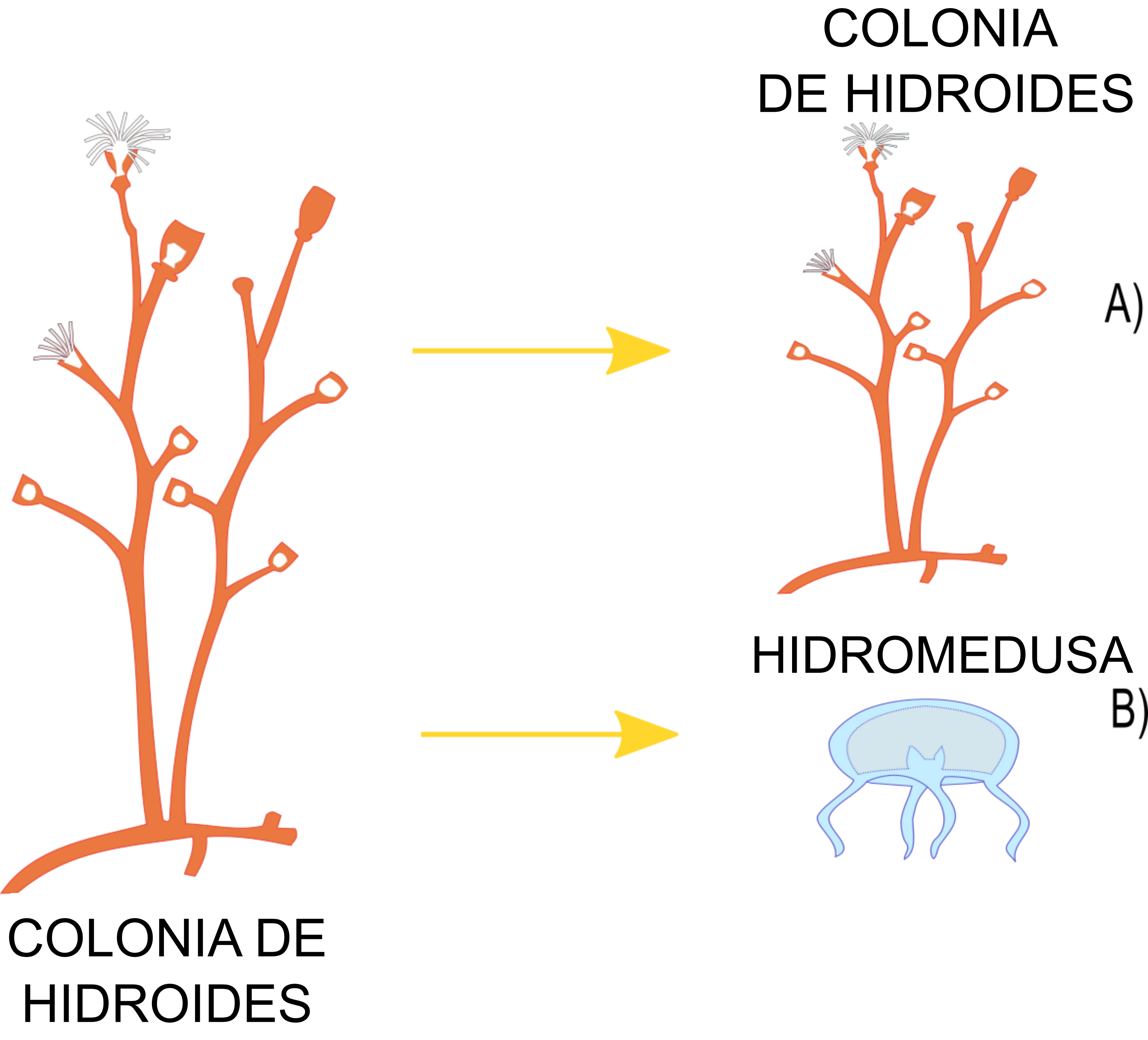
Figura 1. A) Los hidrozoos se pueden reproducir y generar hidroides o B) medusas.arrieras.
Crédito: C. Odette Carral-Murrieta.
La mayoría de los hidroides se mantienen fijos a sustratos, sin embargo, se conocen especies que pueden sobrevivir suspendidas en la columna de agua (por ejemplo, Clytia gracilis; Madin et al., 1996). También hay especies de hidroides que pueden vivir solitarias o formar colonias, es decir que se componen de un conjunto de individuos. En una colonia cada uno tiene una función en particular: algunos cazan o defienden (dactilozoides), otros digieren la comida (gastrozoides) y otros se reproducen (gonozoides) (Mills et al., 2007).
La formación de colonias es común en la mayor parte de las especies de hidrozoos (aproximadamente en el 60%) y está relacionada con su historia evolutiva. Los científicos han considerado que en animales marinos una ventaja de la colonialidad es el aumento de espacio que logran abarcar en las superficies en las que se establecen (Jackson, 1977). Asimismo, cuando un individuo se separa de la colonia, éste puede dar lugar a una colonia nueva, gracias la gran capacidad de regeneración que tienen estos organismos (Gili y Hughes, 1995).
Morfológicamente los hidroides tiene tres características principales: el hidrante, en el cual se encuentra una boca rodeada de tentáculos que le ayudan al animal a capturar presas para comerlas (ver figura 2). El hidrocaule en colonias erectas o pedicelo en colonias estolonales son estructuras parecidas al tallo de las plantas y sirven para mantener de pie al individuo o a la colonia. Por último, pero no menos importante, en los hidroides coloniales tenemos la hidrorriza, la cual es la “raíz” del hidroide, que parece una red y permite al hidroide fijarse en gran variedad de sustratos (Mills et al., 2007; ver figuras 2 y 3).
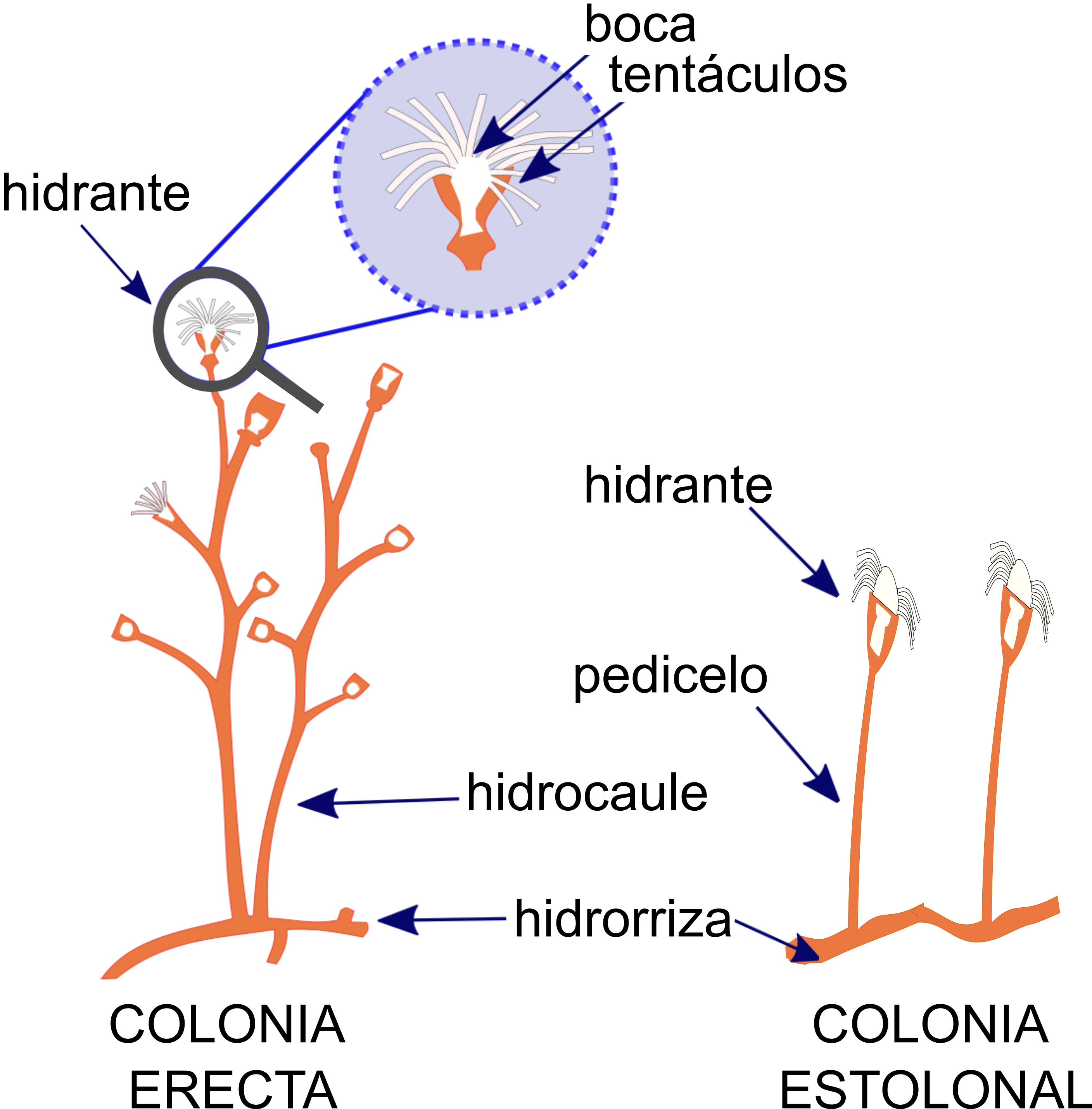
Figura 2. Apariencia general de los hidroides coloniales. El hidroide se compone principalmente del hidrante (con boca y tentáculos), el hidrocaule o pedicelo y la hidrorriza.
Crédito: C. Odette Carral-Murrieta.
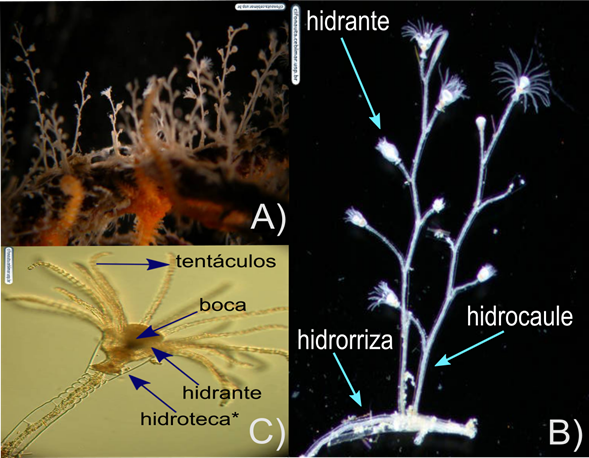
Figura 3. Hidrozoo Obelia dichotoma. A) Colonia. B) Hidrocaule, C) Hidrante. Colonias de hasta 25 mm de altura. *La hidroteca es el exoesqueleto que cubre al hidrante.
Crédito: Alvaro E. Migotto, Cifonauta.
¿Cómo logran los hidroides vivir en tantas superficies?
Es gracias a la hidrorriza que podemos encontrar a los hidroides viviendo sobre otros seres vivos (por ejemplo, ascidias, briozoos, cirrípedos, esponjas, moluscos, poliquetos, otros cnidarios, macroalgas y pastos marinos) o incluso sobre objetos inanimados y construcciones (troncos flotantes, embarcaciones, puertos, muelles, boyas, cuerdas, hilo de pesca, entre otros). Es sabido que eventos naturales como tsunamis pueden arrastrar objetos artificiales por largas distancias, viajando sobre ellos los hidroides, así como permitiendo la fijación de otros animales (Calder et al., 2014).
Una característica maravillosa de la hidrorriza es que, cuando las condiciones del ambiente no son favorables para que el hidroide sobreviva, puede absorber a las demás estructuras del hidroide (hidrante e hidrocaule) y entrar en un estado de latencia en la que el organismo se mantiene vivo hasta que las condiciones ambientales sean favorables y se puedan regenerar las estructuras absorbidas y continuar con su desarrollo (Piraino et al., 2004). Un ejemplo es la especie Clytia hummelincki estudiada en el Mediterráneo, la cual tiene una abundancia máxima entre los meses de agosto y septiembre, mientras que durante la temporada fría las colonias se reducen a tejido latente por medio de la hidrorriza (Martell et al., 2017).
¿Qué tan lejos pueden llegar los hidroides?
Desde la antigüedad, han existido rutas comerciales y exploratorias marítimas entre territorios lejanos. Como ejemplos están la ruta del incienso (siglos iii a. e. c. C. – ii e. c.), que consistía en la navegación dentro del mar rojo, transportando dicho bien entre Asia, África, y hasta terminar en la cuenca del mediterráneo (unesco, 2023); la ruta del galeón de Manila (1565-1815), que unía al puerto de Acapulco con la capital filipina (Martínez-Shaw, 2019). Asimismo, está la famosa expedición de Magallanes, que le dio la vuelta al mundo, saliendo de Europa y pasando por África, el sur de América, Oceanía y Asia (Osada-García, 2022). Por tanto, es muy probable que los seres humanos hayamos comenzado a transportar hidrozoos de manera incidental y por distancias intercontinentales desde hace siglos.
Pero no sólo los barcos pueden recorrer distancias largas, también organismos vivos como las algas pueden llevar a los hidroides a sitios lejanos. Por ejemplo, el alga roja (Acanthophora spicifera), así como las algas cafés llamadas sargazo (Sargassum spp.). Algunas especies de sargazo pasan toda su vida flotando en la superficie del océano, mientras que otras se encuentran fijas a un sustrato, pero se desprenden y quedan a la deriva en el mar (Gutow et al., 2015). Sobre de estos sargazos se fijan varias especies de hidroides que se transportan sobre de ellos, llegando en ocasiones a vararse en las playas (ver figura 4). Hasta el momento, se han reportado 130 especies de hidroides epibiontes en 26 especies de Sargassum1 (Carral-Murrieta et al., 2023).
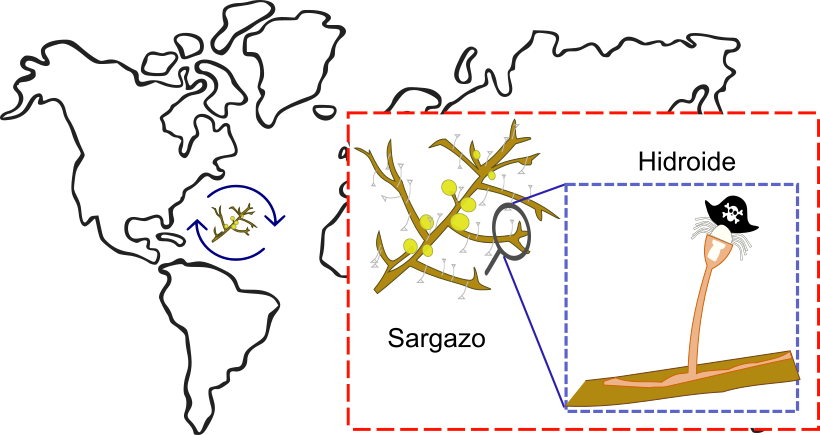
Figura 4. Colonias de hidroides que viajan en el Océano Atlántico sobre el alga café Sargassum spp.
Crédito: C. Odette Carral-Murrieta.
El tiempo de vida de los hidroides que crecen sobre sustratos vivos depende de factores ambientales y del tiempo de vida de los sustratos, así como de la capacidadad de latencia de los hidroides y las interacciones bioticas a las que estén sometidos tanto hidroides como sustratos, por ejemplo, la depredación. En la actualidad se tiene evidencia que permanecen con vida sobre el sargazo que arriba a las playas aun cuando éste queda expuesto a la desecación y descomposición por horas.
Posibles efectos de los hidroides que viajan largas distancias
Los hidroides que llegan a un sitio completamente nuevo se conocen como hidroides exóticos. Algunos de éstos compiten con las especies que viven en el sitio (especies nativas) por espacios para vivir y por las fuentes de alimento. Cuando éstos ocasionan daños y logran establecerse y vivir en el nuevo sitio, se les conoce como hidroides invasores. Sin embargo, los hidroides invasores no sólo impactan a especies nativas, sino que también pueden afectar actividades importantes para nosotros los humanos, tales como el cultivo de animales acuáticos para nuestro consumo, ya que los hidroides pueden ocasionar la muerte de peces; también pueden obstruir tubería importante para el funcionamiento de las plantas de energía, ocasionando pérdidas de tiempo y de dinero (Fitridge et al., 2012).
Actualmente, los científicos han producido algunos métodos para controlar el exceso de organismos como los hidroides viviendo en sustratos artificiales, para así evitar que los seres humanos continuemos introduciendo accidentalmente a especies de hidroides exóticos en sitios en los que previamente no se encontraban. Entre los métodos utilizados para disminuir el impacto de hidroides exóticos que pueden volverse invasores son la creación de pinturas especiales, las cuales evitan que la hidrorriza se fije a los barcos, o tratamientos termales, que modifican la temperatura del agua, lo que previene que los hidroides vivan en zonas en las que se encuentran las superficies que se quieren proteger (Fitridge et al., 2012; Folino-Rore y Indelicato, 2005). Sin embargo, estas pinturas pueden contaminar y ser absorbidas por invertebrados marinos (Turner, 2010). Además, aún no se sabe mucho de los hidroides ni el número de especies que son potencialmente invasoras, por lo que es necesario continuar con la investigación de estos pequeños animales acuáticos.
Conclusión
Los hidroides son animales particulares por su riqueza de especies y ciclos de vida, los cuales están caracterizados por la alternancia de generaciones. Son organismos capaces de adaptarse al medio en el que viven, ya que pueden entrar en latencia cuando las condiciones ambientales son adversas. Además, la hidrorriza les confiere la capacidad de viajar largas distancias sobre una diversidad de superficies flotantes, tales como las algas pardas del género Sargassum. Sin embargo, aún hacen falta más estudios que desentrañen la vida y efectos en los distintos sitios en los que viven.
Agradecimientos
A los proyectos fordecyt-pronaces/428225/2019 “Macroalgas introducidas: organismos modelo para analizar los patrones de éxito de especies con potencial de invasión en ambientes costeros” al Laboratorio de Macroalgas y al equipo de Medusozoa México.
Referencias
- Calder, D. R., Choong, H. H., Carlton, J. T., Chapman, J. W., Miller, J. A., y Geller, J. (2014). Hydroids (Cnidaria: Hydrozoa) from Japanese tsunami marine debris washing ashore in the northwestern United States. Aquatic Invasions, 9(4), 425-440. http://dx.doi.org/10.3391/ai.2014.9.4.02
- Carral-Murrieta, C. O., Marques, A. C., Serviere-Zaragoza, E., Estrada-González, M. C., Cunha, A. F., Fernandez, M. O., Mazariegos-Villarreal A., León-Cisneros, K., López-Vivas, J., Agüero, J. y Mendoza-Becerril, M. A. (En prensa). A survey of epibiont hydrozoans on Sargassum. PeerJ.
- De la Osada García, J. (2022). La ruta de Magallanes-Elcano y el comercio de las especias en España. Anales de la Real Academia de Doctores, 7(2), 329-338. Real Academia de Doctores de España. https://tinyurl.com/2su27reb
- Fitridge, I., Dempster, T., Guenther, J. y De Nys, R. (2012). The impact and control of biofouling in marine aquaculture: a review. Biofouling, 28(7), 649-669. https://doi.org/10.1080/08927014.2012.700478
- Folino-Rorem, N.C. y Indelicato, J. (2005). Controlling biofouling caused by the colonial hydroid Cordylophora caspia. Water Research, 39(12), 2731-2737. https://doi.org/10.1016/j.watres.2005.04.053
- Gili, J. M. y Hughes, R.G. (1995). The ecology of marine benthic hydroids. Oceanography and Marine Biology: An Annual Review, 33, 351-426.
- Global Biodiversity Information Facility (gbif). (2019). Obelia dichotoma (Linnaeus, 1758). https://www.gbif.org/species/5185976
- Gutow, L., Beermann, J., Buschbaum, C., Rivadeneira, M. M. y Thiel, M. (2015). Castaways can’t be choosers – Homogenization of rafting assemblages on floating seaweeds. Journal of Sea Research, 95, 161-171. http://dx.doi.org/10.1016/j.seares.2014.07.005
- Jackson, J. B. C. (1977). Competition on marine hard substrata: the adaptive significance of solitary and colonial strategies. The American Naturalist, 111(980), 743-767.
- Madin, L. P., Bollens, S. M., Horgan, E., Butler, M., Runge, J., Sullivan, B. K., Klein-Macphee G., Durbin E., Durbin A. G., Keuren D. V., Plourde S., Bucklint, A. y Clarke, M. E. (1996). Voracious planktonic hydroids: unexpected predatory impact on a coastal marine ecosystem. Deep Sea Research Part ii: Topical Studies in Oceanography, 43(7-8), 1823-1829.
- Marroig, R. G. y Reis, R. P. (2011). Does biofouling influence Kappaphycus alvarezii (Doty) Doty ex Silva farming production in Brazil? Journal of Applied Phycology, 23(5), 925-931. https://doi.org/10.1007/s10811-010-9602-y
- Martell, L., Ciavolino, E., Gravili, C., Piraino, S. y Boero, F. (2017). Population dynamics of the non-indigenous hydrozoan Clytia hummelincki (Hydrozoa: Campanulariidae) in two contrasting Mediterranean habitats. Marine Biology Research, 13(5), 551-559. https://doi.org/10.1080/17451000.2017.1296160
- Martell, L., Bracale, R., Carrion, S. A., Purcell, J. E., Lezzi, M., Gravili, C., Piraino, F. B. y Boero, F. (2018). Successional dynamics of marine fouling hydroids (Cnidaria: Hydrozoa) at a finfish aquaculture facility in the Mediterranean Sea. PLoS One, 13(4), e0195352. https://doi.org/10.1371/journal.pone.0195352
- Martínez-Shaw, C. (2019). El Galeón de Manila: 250 años de intercambios. Estudis: Revista de historia moderna, 45, 9-34.
- Migotto, A. E. (2007). Hidróide. Colony on the soft coral Leptogorgia punicea. [Fotografía 3A]. Cifonauta. http://cifonauta.cebimar.usp.br/media/1631/
- Migotto, A. E. (s/f). Hidróide. Colony. [Fotografía 3B]. Cifonauta. http://cifonauta.cebimar.usp.br/media/1645/
- Migotto, A. E. (s/f). Hidróide. Hydranth. [Fotografía 3C]. Cifonauta. http://cifonauta.cebimar.usp.br/media/1654/
- Millard, N. A. H. (1975). Monograph on the Hydroida of southern Africa. Annals of the South African Museum, 68, 1-513. https://tinyurl.com/2sz969au
- Mills, C. E., Calder, D. R., Marques, A. C., Migotto, A. E., Haddock, S. H. D., Dunn, C. W. y Pugh, P. R. (2007). Combined species list of hydroids, hydromedusae, and siphonophores. En J. T. Carlton (Ed.), The Light and Smith Manual. Intertidal invertebrates from central California to Oregon (pp. 151-168). University of California Press.
- Turner, A. (2010). Marine pollution from antifouling paint particles. Marine pollution bulletin, 60(2), 159-171. Annals of the South African Museum, 68, 1-513. https://doi.org/10.1016/j.marpolbul.2009.12.004
- Organización de las Naciones Unidas (unesco). (2023). Ruta del incienso – Ciudades del desierto del Neguev. https://whc.unesco.org/es/list/1107
- Piraino, S., De Vito, D., Schmich, J., Bouillon, J. y Boero, F. (2004). Reverse development in Cnidaria. Canadian Journal of Zoology, 82(11), 1748-1754. https://doi.org/10.1139/z04-174
- WoRMS. (2023). World Register of Marine Species. https://www.marinespecies.org